1c)
Komentář: GAG jsou polymery disacharidových jednotek, jejichž základem jsou aminocukr a uronová kyselina. Odpovědi 1a),b),d),e) jsou chybné.
2 b), e)
Komentář: Leucin je ketogenní aminokyselina (degradací neposkytuje pyruvát ani meziprodukty citrátového cyklu), vyšší mastné kyseliny nejsou substráty pro glukoneogenesi. Odpovědi 2a), c), d) jsou chybné – laktát a alanin se přeměňují na pyruvát, glycerol vstupuje do glukoneogenese po fosforylaci na glycerol-3-fosfát a dehydrogenaci na intermediát glukoneogenese dihydroxyacetonfosfát.
3 a), e)
Komentář: za hladovění v důsledku nízkého přísunu glukosy klesá sekrece insulinu, stoupá hladina glukagonu. Glukagon aktivuje hormon-senzitivní lipasu v adipocytech, což vede ke zvýšené lipolýze triacylglycerolů v tukové tkáni a uvolňování mastných kyselin do cirkulace. Odpovědi 3b), c), d) jsou chybné – chylomikrony se tvoří v enterocytu z exogenních lipidů po jídle, za hladovění se tedy v krvi nevyskytují.
4 d), e)
Komentář: Reakce d) představuje inaktivaci noradrenalinu účinkem COMT, reakce e) je první reakcí biosyntézy katecholaminů. Odpovědi 4a), b), c) jsou chybné – ad a) dopamin vzniká z DOPA, ad b),c) produkty reakcí nevznikají bezprostředně z uvedených výchozích látek (přeměna zahrnuje více kroků)
5a), d)
Komentář: GAG jsou součástí proteoglykanů, díky vysokému obsahu polárních a záporně nabitých skupin (OH sacharidových jednotek, karboxylové skupiny uronové kyseliny, sulfátové skupiny) váží vodu (hlavně v důsledku interakce s hydratovanými Na+). Odpovědi 5b),c),e) jsou chybné – běžnou složkou GAG jsou acetylované aminocukry, celkový náboj je záporný (obsah COO-, SO3-), jako antigenní determinanty slouží glykoproteiny.
6d), e)
Komentář: Insulin se váže na insulinový (tyrosinkinasový) receptor, na přenosu signálu v tyrosinkinasové kaskádě se G protein nepodílí. V procesu vidění dochází po interakci světelného kvanta s rhodopsinem k aktivaci membránového G-proteinu (transducinu), následně k aktivaci fosfodiesetrasy a zvýšenému štěpení intracelulárního cGMP, jehož pokles vede k uzávěru kanálu pro Na+ . Odpovědi 6a), b),c) jsou správná tvrzení.
7b), c)
Komentář: glukagon aktivuje fruktosa-2,6-fosfatasu, což vede k defosforylaci fruktosa-2,6- bisfosfátu (alosterického aktivátoru fosfofruktokinasy) – výsledkem je potlačení glykolýzy a stimulace glukoneogenese; Odpovědi 7a), d),e jsou nesprávná tvrzení: prostřednictvím signální kaskády navozuje insulin defosforylaci, glukagon fosforylaci klíčových enzymů metabolismu – proto tvrzení a),d) jsou chybná; insulinem je aktivována acetyl-CoA-karboxylasa, tím tvorba malonyl-CoA (klíčového metabolitu biosyntézy mastných kyselin) a tedy i biosyntéza mastných kyselin.
1.
a)
CH3-CH(NH2)-COO-
+ -OOC-CH2-CH2
–CO-COO- ![]()
alanin 2-oxoglutarát
![]() CH3-CO-COO-
+
-OOC-CH2-CH2–CH(NH2)-COO-
CH3-CO-COO-
+
-OOC-CH2-CH2–CH(NH2)-COO-
pyruvát glutamát
Pozn.: Ve vzorcích mohou být použity i neionizované formy kyselin, i název 2-ketoglutarát, a-ketoglutarát nebo a-oxoglutarát.
a) Patří k enzymovým markerům jaterního poškození. Při poškozeních (např. hepatitida, cirhóza, infekční mononukleóza, intoxikace) jsou v séru zvýšené hladiny tohoto enzymu.
b) 0,15 – 1,2 µkat.l-1
Komentář: U = 60 µkat, tedy převod zadaných hodnot: 9/60 - 72/60
2.
a) Polypeptid - 51 aminokyselin, tvořený 2 řetězci (a a b), které jsou spojeny disulfidovými vazbami. Tvoří se v b-buňkách Langerhansových ostrůvků pankreatu z preproinsulinu (1 řetězec); po odštěpení signální sekvence (v cisternách ER) a vytvoření disulfisulfidových vazeb vzniká proinsulin, z něj po vyštěpení části peptidického řetězce (Golgiho aparát) insulin (ukládán do sekrečních granul).
b) Peptid uvolňující se při přeměně proinsulinu na insulin, skladovaný a sekretovaný spolu s insulinem. Stanovení C-peptidu v plasmě se využívá k diferenciální diagnostice Diabetes mellitus – umožňuje kvantifikovat schopnost pankreatu sekretovat insulin a posoudit typ diabetu.
c) Tuková tkáň patří mezi insulin-dependentní tkáně. V adipocytu insulin stimuluje translokaci glukosových přenašečů (transportérů, GLUT4) z intracelulární hotovosti (zásoby) do plasmatické membrány, tím dochází ke zvýšení počtu přenašečů (pro usnadněnou difuzi glukosy) v membráně a zvýšenému vstupu glukosy do buněk.
3.
a) Apolipoproteiny,
triacylglyceroly, cholesterol volný i esterifikovaný, fosfolipidy
(vysoký obsah apolipoproteinů
a fosfolipidů, nízký obsah TAG).
b) Místo tvorby - játra;
funkce – reverzní transport cholesterolu z tkání do jater
zásobárna apoplipoproteinů pro ostatními lipoproteiny (zejména Apo C a E pro VLDL a chylomikrony)
c) LCAT (lecithincholesterolacyltransferasa) katalyzuje přenos mastné kyseliny z fosfolipidů na cholesterol v povrchu částice HDL – význam pro mechanismus reverzního transportu cholesterolu.
Pozn: na otázky lze odpovídat heslovitě, viz tučně vyznačené části textu.
1. Doplněné schema (červeně):
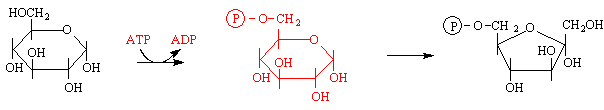
1 glukosa-6-fosfát fruktosa-6-fosfát
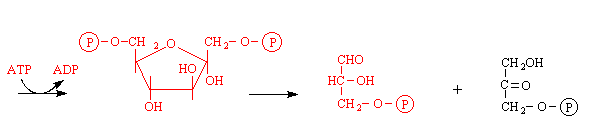
2 fruktosa-1,6-bisfosfát 3 glyceraldehyd-3-fosfát dihydroxyacetonfosfát
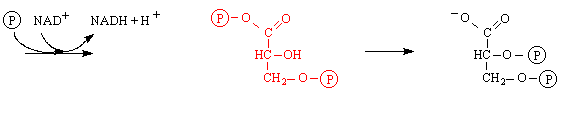
4 1,3-bisfosfoglycerát 5 2,3-bisfosfoglycerát
2.
- cytosol
- erythrocyt; regulace afinity kyslíku k hemoglobinu
- nedostatečná saturace periferních tkání kyslíkem, např. pobyt ve vyšších nadmořských výškách
3.
- glukokinasa, hexokinasa
- glukokinasa – játra (buňky pankreatických ostrůvků), hexokinasa – ostatní tkáně
- Km, t.j. afinitou ke glukose (glukokinasa má nízkou afinitu, hexokinasa vysokou)
-
vzhledem k vysoké Km (nízké
afinitě) glukokinasy pro glukosu je v játrech metabolizována
jen část vstupující glukosy, zbylá část přechází do cirkulace;
v extrahepatálních
tkáních pak zajišťuje hexokinasa (díky nízké Km
,vysoké afinitě) účinnou
dodávku glukosy buňkám (i při nízkých hladinách glukosy v krvi).